1911年にラモニ・カハール(1906年ノーベル医学生理学賞受賞)らによって海馬の主要な記憶神経回路である、トライシナプス性(嗅内皮質→ DG(歯状回)→CA3→CA1) の記憶神経回路が初めて発見されました。
現在までのおよそ100年間、トライシナプス性の記憶神経回路は、最も有名で重要な記憶神経回路の1つとされています。
研究内容Research
記憶は、私たちの日常生活の一部となっている脳機能です。
しかし、記憶のメカニズムは部分的にしか解明されておらず、自然科学に残された最大スケールの謎のひとつであります 。私は、新規戦略技術を用いて、分子、神経回路、局所領域、行動といった総合的なアプローチで記憶研究を行ってきました。
私たちは、遺伝子組換えウィルス、分子生物学、光遺伝学、パッチクランプ法などの電気生理学、遺伝子組み換えマウスを用いて、記憶神経回路、新CA2領域、新海馬地図と社会性記憶のメカニズムおよび「遺伝子の戦い」を利用した新戦略技術「BATTLE」や新たな戦略技術開発を中心に研究を展開しています。
これまでの研究概要
研究1.「新CA2領域と新海馬地図」

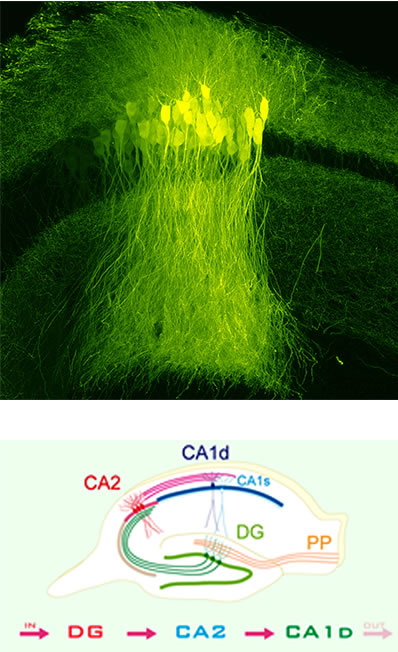
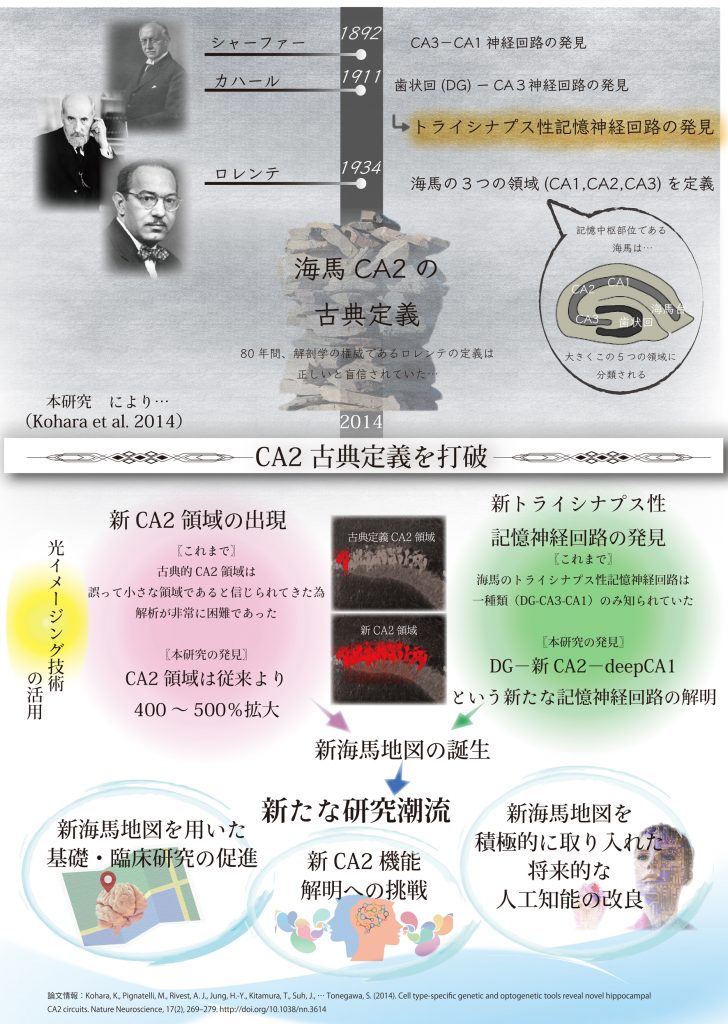
一方、CA2は、1934年にロレンテ・デノによって定義されたが、解明が最も遅れていました。
私たちは、先端複合光技術を用い、80年以上にわたり信じられていた古典定義を覆し、CA2がこれまで考えられていたよりもはるかに大きい新CA2領域であることを世界に先駆けて証明し、これにより新CA2領域が出現しました。さらに、私たちは新規のトライシナプス性の記憶神経回路を発見しました。
(Kohara K# et al. Nat. Neuroscience, 2014, #筆頭代表著者)
これにより、記憶中枢部位海馬において、突如として事実上の新規研究可能領域(新CA2領域)と新海馬地図が誕生しました。
- この新しい研究潮流に、現在ノーベル賞受賞者を含む多くの有力研究者たちが続々と新規参入しています。
- ( Dudek et al., Nat. Rev. Neurosci 2016
Whalley et al., Nat. Rev. Neurosci 2016
Lewis et al., Nat. Rev. Neurosci 2014
Bayer et al., Nat. Neurosci 2014
Hitti, et al., Nature 2014
Rajasethupathy et al., Nature 2015
Kay et al., Nature 2016
Leroy et al., Nature 2018
Oliva et al., Nature 2020
Sun et al., Nature 2020
Kitamura et al., Science 2014
Suh et al., Cell 2019
Kay et al., Cell 2020
Valero et al., Nat. Neurosci 2015
Li et al., Nat. Neurosci 2017
Donegan et al., Nat. Neurosci 2020
Mankin et al., Neuron 2015
Lee et al., Neuron 2015
Miao et al., Neuron 2015
Lu et al., Neuron 2015
Piscorowski et al., Neuron 2016
Oliva et al., Neuron 2016
Boehringer et al., Neuron 2017
Cembrowski et al., Neuron 2017
Leroy et al., Neuron 2017
Sun et al., Neuron 2017
Fernández-Ruiz et al., Neuron 2017
Tirko et al., Neuron 2018
Roussarie et al., Neuron 2020
Alexander et al., Nat. Comm 2016
Meira et al., Nat. Comm 2018
Raam et al., Nat. Comm 2018
Domínguez et al., Cell Rep 2019
Farris et al., Cell Rep 2019
Nasrallah., Cell Rep 2019
Fernandez-Lamo et al., Cell Rep 2019
Sun et al., eLife 2014
Wheeler et al., eLife 2015
Cembrowski et al., eLife 2016
Alexander et al., eLife 2018
Pagani et al., Mol Psychiatry 2015
Smith et al., Mol Psychiatry 2016
Wintzer et al., J. Neurosci 2014
Mitre et al., J. Neurosci 2015
Carstens et al., J. Neurosci 2016
Williams et al., PLOS genetics 2016
Stevenson,et al Eur J Neurosci. 2014
Botcher et al., Front Neuroanat. 2014
Llorens-Martín Exp Neurol. 2015
Brown et al., J Biol Chem 2015
Nasrallah et al., eNeuro 2015
Choi et al., J. veterinary Science 2015
Liao et al., Mol. Medicine reports 2015
Nunes et al., PLOS one 2015
Häussler et al., Hippocampus 2015
Hisamatsu et al., Neuroreport 2016
Lee et al., J Neuroinflamation 2016
Cao et al., Neuroscience Letters 2016
Lee et al., Neuroscience 2015
Faraji et al., Behav Brain Res 2016 他多数)
研究2. Cre recombinase を軸とした新研究戦略・技術開発
現代科学における大きな2本柱として「新研究技術・ツール」と「新発見」があります。
これらの2つの柱は、「ニワトリと卵の関係」にあるといえます。
すなわち、新研究技術・ツールによって新発見が生み出され、また一方で、新発見によって新研究技術・ツールが生み出されます。私たちは、主に遺伝子組換え酵素Cre リコンビナーゼを軸とした新研究戦略・技術開発を行なってきました。
1.遺伝子の戦いを活用した新研究戦略・技術開発
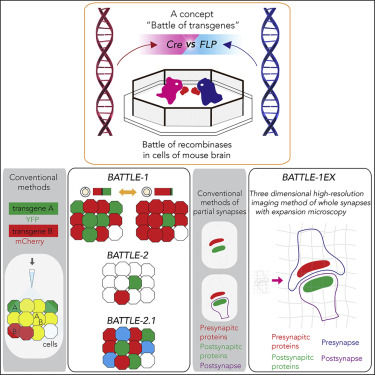
枚方の関西医科大学の芝生の小道を歩いている際に、「遺伝子同士を戦わせたらどうなるか?」という着想を独自に得ました。それを発展させ「遺伝子の戦い」という概念を発表しました。そして、その概念に基づいて、遺伝子組換え酵素同士を戦い合わせる戦略技術群「BATTLE」を新規開発しました。
「BATTLE-1」は、これまで困難であった自在な反発分離的遺伝子導入の実現のための新技術、「BATTLE-2」は、複数の遺伝子のまばらな遺伝子導入の実現のための新技術、そして「BATTLE-2.1」は、3種類の遺伝子の反発分離的遺伝子導入の実現のための新技術です。
これらの「BATTLE」技術を用いて、マウス脳内の海馬で人工的に「遺伝子の戦い」を起こさせることに成功しました。
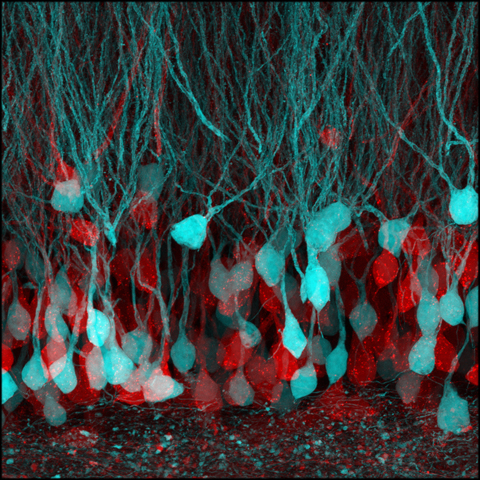
脳神経回路の接合部であるシナプスは、脳の神経活動の要ともいえる根本的な部位です。しかしながら、シナプスは複数の神経細胞からなる構造であることから、シナプスの全体像を高精細に可視化することは、これまでの光を用いた既存技術では困難でした。
私たちは、「BATTLE」技術を既存の膨張顕微鏡技術と融合させた複合技術「BATTLE-1EX」を開発し、マウス海馬の神経回路の接合部であるシナプスに対して、プレシナプス、ポストシナプスの構造および、シナプス構成タンパク質の局在も含めて、シナプスの全体像を高精細に可視化することに成功しました。
2. 1細胞遺伝子ノックアウト戦略技術の開発
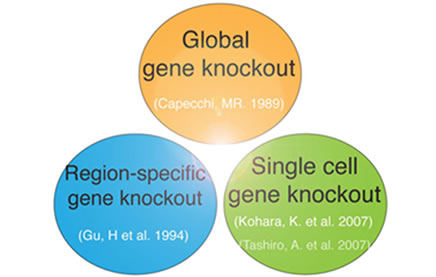
遺伝子の機能を調べる手法である遺伝子ノックアウト技術は、ノーベル賞が授与された王道の中の王道の実験技術です。その当時は、全身や特定領域の大多数の細胞において遺伝子ノックアウトし機能を調べるスタイルが全盛でした。
私たちは、Cre recombinase を単一細胞に遺伝子導入することで、この方向性とは真逆の、究極的に少数の1細胞で遺伝子ノックアウトするという全く新たな価値観に基づく第三世代の実験方法戦略を開発しました。(Kohara K. et al. J. Neurosci, 2007)
- この戦略技術は、その後、医学生物学、記憶研究分野において世界標準的に用いられる戦略技術の一つとなっています。
- (Lin Y et al. Nature 2008, Adesnik et al., PNAS 2008, Lu W et al. Neuron 2009, Lu et al., PNAS 2010, Odajima et al., Dev Cell 2011, Lu et al., Neuron 2013, Herring et al., Neuron 2013, Granger AJ et al. Nature 2013, Bloodgood BL et al. Nature 2013, Lovero et al., PNAS 2015, Levy et al., Neuron 2015, Mizuno et al Neuron 2016他多数)
研究3.「記憶形成分子BDNFに関する研究」
脳由来神経栄養因子BDNFは、記憶学習において必須な物質であり、脳科学分野において最も注目される分子の一つです。当時、非常に強固な定説があり、ほとんどすべての研究者が「BDNFはシナプスの後部から前部へと逆行性に作用する。」と信じていました。
私たちは、BDNF-GFPが神経活動に依存してシナプス前部からシナプス後部へ順行性に移行することを発見しました。
(Kohara K. et al. Science, 2001)
さらに新規の実験技術であるキメラ培養法を開発することによって、内在性のBDNFが順行性に移行することを発見しました。
(Kohara K. et al. Journal of Neuroscience, 2003)
これにより、BDNFがシナプス後部からだけでなくシナプス前部からも作用していることが明らかになり、記憶形成分子の新たな作用経路が発見されました。
その後、多くの研究グループによる研究も加わり 、長年世界で信じられてきた定説は書き換えられました。
(脳科学辞典「神経栄養因子」小原圭吾)
(Anastasia et al., Nat. Rev. Neurosci, 2013, Park et al., Nat. Rev. Neurosci, 2013,他多数)